
-
![画像]()
渡り蝶2種の標本。左上:アサギマダラ♂、表面、神奈川、右上:アサギマダラ♂、裏面、奄美大島(長野→奄美大島の約1,200 kmの移動が確認されたマーキング個体・芦澤一郎採集)、左下: オオカバマダラ♂、表面、オーストラリア、左下: オオカバマダラ♀、表面、ハワイ
-
![画像]()
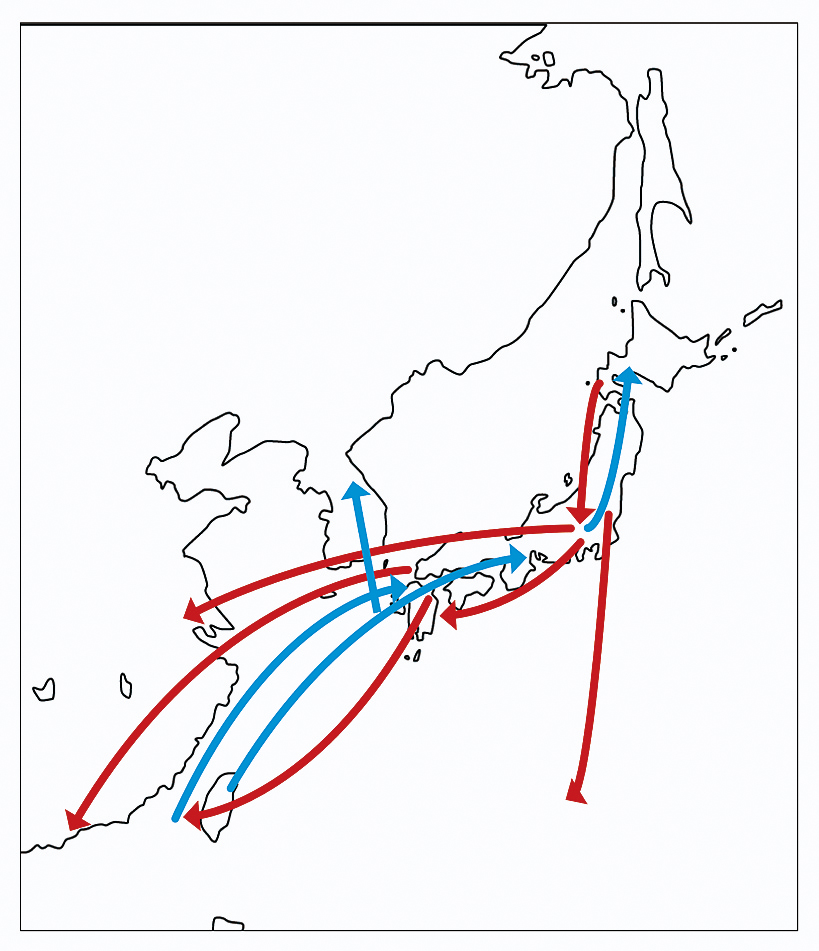
アサギマダラの長距離移動の例。金沢ほか(2013)を改変、青線は春〜夏にかけての北上移動、赤線は晩夏〜秋にかけての南下移動を示す
-
![画像]()
北米産オオカバマダラの集団越冬(撮影:倉地 正)


B25
アサギマダラとオオカバマダラ
渡りをする蝶
周期的かつ季節的に一定方向を旅する「渡り」が知られる蝶として、日本ではアサギマダラ、北米ではオオカバマダラがいる。両種ともタテハチョウ科・マダラチョウ亜科に属し、有毒のガガイモ科植物を幼虫が食べることから、成虫も食草由来の有毒成分を体内に蓄積するため、鳥などの天敵から捕食されにくいと考えられている。
アサギマダラの渡りに関しては、1980年頃から翅に標識をつけて放し(マーキング)、それを再捕獲する市民参加型の調査が各地でなされ、春〜夏には台湾・南西諸島から数世代で本州・北海道へ北上し、秋には逆のコースで一気に北海道・本州から南西諸島・台湾、時には中国大陸まで一世代で南下することが判明し、新聞やテレビ等でもよく取り上げられている。2011年には和歌山から高知を経由して香港まで約2,500kmを移動した個体が再捕獲されて話題となり、これが現在の最長移動記録となっている。また、本種の約6亜種でそれぞれ移動性の有無が異なり、最近の分子系統学的研究によると、本種の祖先は定住性で、一部の個体群(例えば日本亜種)が移動性に派生したと考えられている。各亜種レベルでの遺伝的距離も大きく、複数の隠蔽種が本種内に混在する可能性が高い。この結果は成虫の斑紋や性標、幼生期のデータからも支持されている。
一方、北米産のオオカバマダラは春から夏に3〜4世代を重ねて北上してアメリカ北部やカナダに達し、秋に北米北部からカリフォルニア南部やメキシコまで3,000km以上も一気に南下した後、樹上の枝や葉などに止まりながら集団で越冬する。渡りの機構については、太陽コンパスや磁気コンパスが体内に存在し、これにより定位することや、大きな飛翔効率を生む遺伝子なども判明している。また、比較ゲノム解析からオオカバマダラ属は祖先が移動性で、北米から分散して中南米に定住性の種が広く分布するようになったという、アサギマダラとは逆の進化が起こったことが明らかにされている。 (矢後勝也)
参考文献 References
金沢 至・橋本定雄・福村拓己・伊藤雅男・アサギマダラを調べる会(2013)「アサギマダラの移動における日本海ルートの可能性」『Nature Study』60 (1): 3–6。
新川 勉・矢後勝也・中 秀司・福田晴夫・村上 豊・宮武頼夫・野中 勝(2006)「マダラチョウ科の分子系統」『昆虫と自然』42 (1): 5–11。
矢後勝也(2012)「2011年の昆虫界をふりかえって—蝶界」『月刊むし』(495): 2–18。
矢後勝也(2015)「第6章・チョウにみる進化と多様化」『遺伝子から解き明かす昆虫の不思議な世界』大場裕一・大澤省三・昆虫DNA研究会(編):251–310、悠書館。