
小石川分館特別展示
貝殻微細構造
貝殻は生物がつくる鉱物(生体鉱物)の1種である。生体鉱物を持つ生物は非常に多いが、貝類の貝殻の特徴を一言で表現するとすれば、ベニア板のように強化された合板である。微細構造から明かなように、貝殻は陶器のように均質なものではなく、複数の異なる層(殻層)が重なり合ってできている。性質の異なる層を重ね合わせることは、貝殻全体の強度を増す工夫であり、他の生物には成し得ない極めて高度な外骨格の形成様式である。
貝類には大小様々な種が存在する。現生種で最大のものは現生ではオオシャコガイ(二枚貝)で1m以上になる。一方、最小の貝は、成体ではミジンワダチガイ類であり殻径0.6mm程度である。幼生まで入れれば、小さいものはベリジャー幼生の貝殻で0.2mmである。大きい貝も小さい貝も全て複数の殻層からできている。そして、ひとつひとつの殻層は、さらに細かい結晶の集合体でできており、結晶と有機物はレンガとモルタルの関係にたとえられる。
貝殻の主成分は炭酸カルシウムであり、カルシウム、炭素、酸素からなる。炭酸カルシウムには構造的な多型があり、貝殻はアラゴナイト(霰石)とカルサイト(方解石)の2種類の組み合わせからなる。アラゴナイトとカルサイトは密度(重さ)や溶解に対する抵抗性が異なるため、両者の選択は貝類の生存戦略と関係があると考えられる。例えば、殻を同じ厚さで少しでも軽くしたい場合には、カルサイトの比を増やす方が有利になる。
軟体動物のマクロな多様性は、種数が多いこと(種多様性)、形やボディプランが変化に富むこと(形態学的多様性)、幅広い環境に適応すること(生態的多様性)、ゲノムレベルでもグループ間に大きな差が見られること(遺伝的多様性)、化石記録が豊富であること(古生物学的多様性)がある。このような多様性は軟体動物の特徴のひとつであるが、それは結晶レベルでも当てはまる。貝殻の結晶は40種以上のタイプに細分されており、生体鉱物を持つ生物の中では最も複雑な結晶の組み合わせを持つ。
貝殻の内部構造を観察するには、貝殻を破断するか、切断して断面を研磨する(図1)。貝殻の内部には成長線、殻層、結晶の3つの構造がある。成長線は樹木の年輪のように成長の履歴を示す線であり、殻層、微細構造とは独立にできる。殻層は、微細構造が異なる層状の構造単位であり、4層から6層の殻層を持つ種が多い。筋肉が付着する部位は殻筋層と呼ばれる特別な薄い層が形成される。殻層が4層の場合は、殻筋層を除外すると3層になり、外側から外層、中層、内層と呼ばれている(図2)。殻層は地層の重なりのようなもので、殻の層の境界部は不連続面として識別でき、色が異なることが多い(図2)。
それぞれの殻層の内部を拡大すると、殻層を構成する微小な結晶を見ることができる。そして、結晶の形、大きさ、成長面に対する結晶の傾斜角度などの形質の組み合わせにより、微細構造を定義することができる。結晶の形態を正確に観察するためには、殻の表面に平行な方向、殻の表面に垂直な方向で殻の成長軸に平行な方向、殻の表面に垂直かつ殻の成長軸に垂直な方向の3通りの向きから観察するとよい。
最もよく用いられる微細構造の用語は、真珠構造、稜柱構造、交差板構造、葉状構造、均質構造である。真珠構造(図3A-B)は薄い板状の結晶が重なりあって出来ており、真珠も薄板の積層でできている。真珠光沢を持つ種は、真珠構造を持っており、その色が構造色であることは広く知られている。板状の結晶の間には厚い有機質の膜があり、結晶を硬く結びつける役割をしている。真珠構造は最も頑丈な貝殻構造であることが知られているが、有機物を他の微細構造よりも多量に必要とするため、殻の形成コストは高いと考えられる。
稜柱構造(図3C)は柱状の構造である。断面は円形であったり、多角形であったり、不規則であったり、様々であるが、結晶の成長方向に沿って細長く伸びる。稜柱構造は殻の最も外側に位置することが多く、殻の溶解を防ぐ重要な機能を果たしていると考えられる。
交差板構造(図2, 3D)は、隣り合う板状の結晶が互い違いになるように角度を変えながら成長する。となり合う結晶の伸びる角度がなぜ逆になるのか、合理的な説明は存在していない。この構造は有機物の含有量が比較的少ないことが知られており、形成のコストは低いと考えられる。交差板構造を持つ種は巻貝、二枚貝ともに非常に多く、軟体動物において最も重要な構造のひとつである。
葉状構造は、真珠構造によく似た板状の構造であるが、真珠構造がアラゴナイトであるのに対して、葉状構造はカルサイトからなる。二枚貝の翼足類の一部に見られる構造であり、巻貝ではカサガイ類の一部を除いて見られず、貝類全体から見れば葉状構造を持つ種は少数派である。
均質構造は、小さい顆粒状の結晶の集合である。従って、どの面から見ても粒状の結晶が見える。この構造は比較的薄い層であることが多い。
貝殻は異なる微細構造の組み合わせできているが、層の数、配置、微細構造の分布の組み合わせは、分類群間で異なる例が多い。特に、属、科レベルの分類群で特有の微細構造の分布を示す例が多い。このことから、微細構造は比較的低次の分類群の定義形質、分類形質として利用することができる。従って、貝殻の微小な断片さえも、微細構造が保存されていれば分類学的同定が可能であり、化石の研究で用いられてきた。さらに、近年では遺跡から出土した加工品の鑑定など考古学の分野でも微細構造の形質を利用している例がある。
我々にとって理解が難しいことは、貝殻の成長はミクロな結晶レベルの話である一方、巨視的には極めて規則正しい螺旋を作り出しているという複雑な成長様式である。結晶や螺旋を生み出す設計図はゲノム中にどのように保存され、制御されているのか謎である。そもそも貝殻がどのような反応経路を経て形成されているのか、現状では分子レベルでの完全な説明は無く、貝殻は謎に満ちている。
ウロボロスVolume24 Number2のトップページへ
貝殻微細構造
佐々木猛智(本館准教授/動物分類学・古生物学)
貝殻は生物がつくる鉱物(生体鉱物)の1種である。生体鉱物を持つ生物は非常に多いが、貝類の貝殻の特徴を一言で表現するとすれば、ベニア板のように強化された合板である。微細構造から明かなように、貝殻は陶器のように均質なものではなく、複数の異なる層(殻層)が重なり合ってできている。性質の異なる層を重ね合わせることは、貝殻全体の強度を増す工夫であり、他の生物には成し得ない極めて高度な外骨格の形成様式である。
貝類には大小様々な種が存在する。現生種で最大のものは現生ではオオシャコガイ(二枚貝)で1m以上になる。一方、最小の貝は、成体ではミジンワダチガイ類であり殻径0.6mm程度である。幼生まで入れれば、小さいものはベリジャー幼生の貝殻で0.2mmである。大きい貝も小さい貝も全て複数の殻層からできている。そして、ひとつひとつの殻層は、さらに細かい結晶の集合体でできており、結晶と有機物はレンガとモルタルの関係にたとえられる。
貝殻の主成分は炭酸カルシウムであり、カルシウム、炭素、酸素からなる。炭酸カルシウムには構造的な多型があり、貝殻はアラゴナイト(霰石)とカルサイト(方解石)の2種類の組み合わせからなる。アラゴナイトとカルサイトは密度(重さ)や溶解に対する抵抗性が異なるため、両者の選択は貝類の生存戦略と関係があると考えられる。例えば、殻を同じ厚さで少しでも軽くしたい場合には、カルサイトの比を増やす方が有利になる。
軟体動物のマクロな多様性は、種数が多いこと(種多様性)、形やボディプランが変化に富むこと(形態学的多様性)、幅広い環境に適応すること(生態的多様性)、ゲノムレベルでもグループ間に大きな差が見られること(遺伝的多様性)、化石記録が豊富であること(古生物学的多様性)がある。このような多様性は軟体動物の特徴のひとつであるが、それは結晶レベルでも当てはまる。貝殻の結晶は40種以上のタイプに細分されており、生体鉱物を持つ生物の中では最も複雑な結晶の組み合わせを持つ。
貝殻の内部構造を観察するには、貝殻を破断するか、切断して断面を研磨する(図1)。貝殻の内部には成長線、殻層、結晶の3つの構造がある。成長線は樹木の年輪のように成長の履歴を示す線であり、殻層、微細構造とは独立にできる。殻層は、微細構造が異なる層状の構造単位であり、4層から6層の殻層を持つ種が多い。筋肉が付着する部位は殻筋層と呼ばれる特別な薄い層が形成される。殻層が4層の場合は、殻筋層を除外すると3層になり、外側から外層、中層、内層と呼ばれている(図2)。殻層は地層の重なりのようなもので、殻の層の境界部は不連続面として識別でき、色が異なることが多い(図2)。
それぞれの殻層の内部を拡大すると、殻層を構成する微小な結晶を見ることができる。そして、結晶の形、大きさ、成長面に対する結晶の傾斜角度などの形質の組み合わせにより、微細構造を定義することができる。結晶の形態を正確に観察するためには、殻の表面に平行な方向、殻の表面に垂直な方向で殻の成長軸に平行な方向、殻の表面に垂直かつ殻の成長軸に垂直な方向の3通りの向きから観察するとよい。
最もよく用いられる微細構造の用語は、真珠構造、稜柱構造、交差板構造、葉状構造、均質構造である。真珠構造(図3A-B)は薄い板状の結晶が重なりあって出来ており、真珠も薄板の積層でできている。真珠光沢を持つ種は、真珠構造を持っており、その色が構造色であることは広く知られている。板状の結晶の間には厚い有機質の膜があり、結晶を硬く結びつける役割をしている。真珠構造は最も頑丈な貝殻構造であることが知られているが、有機物を他の微細構造よりも多量に必要とするため、殻の形成コストは高いと考えられる。
稜柱構造(図3C)は柱状の構造である。断面は円形であったり、多角形であったり、不規則であったり、様々であるが、結晶の成長方向に沿って細長く伸びる。稜柱構造は殻の最も外側に位置することが多く、殻の溶解を防ぐ重要な機能を果たしていると考えられる。
交差板構造(図2, 3D)は、隣り合う板状の結晶が互い違いになるように角度を変えながら成長する。となり合う結晶の伸びる角度がなぜ逆になるのか、合理的な説明は存在していない。この構造は有機物の含有量が比較的少ないことが知られており、形成のコストは低いと考えられる。交差板構造を持つ種は巻貝、二枚貝ともに非常に多く、軟体動物において最も重要な構造のひとつである。
葉状構造は、真珠構造によく似た板状の構造であるが、真珠構造がアラゴナイトであるのに対して、葉状構造はカルサイトからなる。二枚貝の翼足類の一部に見られる構造であり、巻貝ではカサガイ類の一部を除いて見られず、貝類全体から見れば葉状構造を持つ種は少数派である。
均質構造は、小さい顆粒状の結晶の集合である。従って、どの面から見ても粒状の結晶が見える。この構造は比較的薄い層であることが多い。
貝殻は異なる微細構造の組み合わせできているが、層の数、配置、微細構造の分布の組み合わせは、分類群間で異なる例が多い。特に、属、科レベルの分類群で特有の微細構造の分布を示す例が多い。このことから、微細構造は比較的低次の分類群の定義形質、分類形質として利用することができる。従って、貝殻の微小な断片さえも、微細構造が保存されていれば分類学的同定が可能であり、化石の研究で用いられてきた。さらに、近年では遺跡から出土した加工品の鑑定など考古学の分野でも微細構造の形質を利用している例がある。
我々にとって理解が難しいことは、貝殻の成長はミクロな結晶レベルの話である一方、巨視的には極めて規則正しい螺旋を作り出しているという複雑な成長様式である。結晶や螺旋を生み出す設計図はゲノム中にどのように保存され、制御されているのか謎である。そもそも貝殻がどのような反応経路を経て形成されているのか、現状では分子レベルでの完全な説明は無く、貝殻は謎に満ちている。
ウロボロスVolume24 Number2のトップページへ
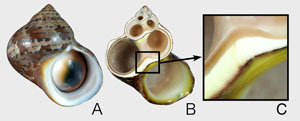
図1 リュウテンサザエ. A. 殻の外観. B. 殻の縦断面. C. 貝殻の切断面に見える殻層.
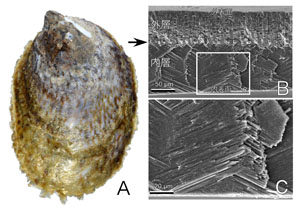
図2 シマメノウフネガイ. A. 殻の外観(背面図). B-C. 走査電子顕微鏡像. B. 殻縁付近での殻の垂直断面. 外層と中層が見えている. 矢印は2つの層の境界. 外層と中層は両者とも交差板構造であるが結晶の成長方向が異なる. C. 交差板構造の拡大図.

図3 貝殻微細構造の走査電子顕微鏡像. A. 真珠構造(シート状真珠構造). ムラサキイガイ. B. 真珠構造(柱状真珠構造). コシダカエビス. C. 稜柱構造(繊維状稜柱構造). ムラサキイガイの内表面. D. 交差板構造. ヒメヨウラク.