
研究紹介
海洋島で種分化したチョウの謎を解く
―小笠原固有のチョウ「オガサワラセセリ」―
本稿の主役・オガサワラセセリは年4~5回程度発生する多化性で、食草オガサワラススキ(イネ科)が繁茂する海岸線の草原帯で発生する(図2)。1905年に北海道帝国大学教授の松村松年(当時は札幌農学校教授)らにより父島で発見され、その翌年に松村自身の手により新種として発表されたのが最初である(Matsumura, 1906)。1935年には母島でも確認され(竹内, 1936b)、戦後になって母島属島の平島、向島、姉島からも記録された(加藤, 1991)。苅部ら(2004)により同じく母島属島の妹島、姪島からも報告がある。ただし父島からは戦前の記録しかない。また、戦後は1970年に母島で再発見されるまで長らく記録されなかった。そのためにオガサワラシジミが国の天然記念物に指定された1969年の時点で本種の生息が確認できず、文化庁による天然記念物選定の名簿から漏れた経緯があるが、生息範囲が狭いことからその希少性は高い。
このような本種であるが、実は最近になって北米原産の外来トカゲ・グリーンアノール(図3)の捕食圧により、これらのチョウ類をはじめとした多くの固有昆虫に絶滅の危機が迫っている(矢後・中村, 2007;矢後, 2014)。1960年年代に資材混入あるいはペットとして侵入したものがまず父島で増殖し、1984年に母島へ移入されて急増、さらに2013年には兄島でも多数繁殖していることが確認された。その生息数は現在500万頭とも600万頭とも推算されている。加えて海岸線ではモクマオウやリュウキュウマツ、ギンネム等の外来植物が、オガサワラセセリの食草オガサワラススキを被陰してその生長を阻害し、チョウ幼虫の成育に甚大な影響を及ぼしている(図4)。このため、環境省は2018年に種の保存法(絶滅のおそれのある野生動植物の種の保存に関する法律)の国内希少野生動植物種に指定し、捕獲・譲渡等を規制するとともに、母島やその属島での生息地の保全や保護増殖事業計画の策定等を行う準備を進めている。
一方、父島では戦後の記録がないことを述べたが、これはなぜだろうか?おそらく戦前の飛行場建設や築港の増加、宅地拡大、農地開発やそれに伴う農薬散布、軍の駐留による要塞化などで発生地のコアエリアが破壊されたことが主要因となり、残念ながら1940年代には絶滅したと考えられる。本種の存続には複数箇所の広大な海岸性草原が必要と思われ、台風や高波などによる塩害や季節的な変化、風向き、日照条件などから、時期ごとや年度ごとに幼生期の最適な生息場所や食草条件が少しずつ異なり、これに合わせて主要な発生場所を移動させているようである。要衝の中心であった父島やその属島では、そのような海岸性草原が真っ先に失われた可能性が高い。 父島からは1905年における本種最初の発見(Matsumura, 1906;松村, 1906)の他、戦前の朝比奈(1932)と竹内(1936b)による記録の計3例しか知られていない。しかも朝比奈の記録は伝聞、竹内の記録は目撃に留まり、これまで標本として存在するのは北海道大学に保管されている松村松年らのわずかな採集品のみであった。ところが、2017年冬に(公財)山階鳥類研究所から東大総合研究博物館に移管された昆虫標本の中から父島産オガサワラセセリの標本(図5)が見出されたのである(矢後ら, 2018)。ラベルには「Nov. 23, 1930. Chichijima, Ogasawara. K. Omichi leg.」と記載されている。標本の採集者K. Omichiは、小笠原島庁の勧業課長や植物検査官補としてミバエ類駆除の調査研究に従事していた大道金松と思われ(石井, 2015)、朝比奈(1932)の文中にも「ヲガサハラセセリ等に至つては島廳の大道氏の御話によると近年全く得られず、僅に同氏が本年の二月とかに二頭を得たに過ぎぬとの御話でした。」の記述が見られる。つまり大道がオガサワラセセリに精通し、実際に採集していたことは疑いない。故にこの標本データの信憑性は極めて高いことから、父島に間違いなく生息していたことを示す極めて重要な標本となるだろう。
ところで、小笠原のような海洋島でいつ、どこから、どのようにオガサワラセセリの祖先種が入り込み、固有種として分化したのか、という疑問が生じる。本種を含むセセリチョウ科のイチモンジセセリ属Parnaraは、アジアからオーストラリア、アフリカに分布し、世界からおよそ10種が知られ、日本にはオガサワラセセリP. ogasawarensisの他に、近縁種として本州以南に生息するイチモンジセセリP. guttataと南西諸島以南に分布するヒメイチモンジセセリP. badaの2種が生息する。特にオガサワラセセリはイチモンジセセリに最近縁とされていたものの、オーストラリアに分布するオセアニアイチモンジセセリP. amaliaに斑紋や♂交尾器がより類似することから、系統的位置の再検討も必要とされてきた。そこで筆者は中国の華南農業大学等との共同研究により、イチモンジセセリ属全種を用いて8つの遺伝子領域に基づいて分子系統解析を行い、分類学的再検討と系統地理学的考察を行った(Huang et al., 2019)。その結果が図6である。本属の共通祖先は漸新世中期に生じ、中新世後期から鮮新世後期にかけてアジア、オーストラリア、アフリカの各地域で分化を起こし、その後の分散と分断がアジアとアフリカでの多様化に大きな役割を果たしたと考えられた。ここで最も注目したいオガサワラセセリの系統的位置は、ヒマラヤ~中国、台湾、日本に広く生息するイチモンジセセリと中国南部~インドシナに分布するカクレイチモンジセセリP. battaで形成されるクレードの姉妹群として表されている。小笠原では稀にイチモンジセセリが記録されるが、オガサワラセセリはイチモンジセセリとは姉妹種にならないため、オガサワラセセリが独立種として維持されていることが理解できる。さらにオガサワラセセリの祖先種は北の日本本土方面や南のオセアニア方面の経由ではなく、むしろ琉球弧形成前の南西方面を経由して鮮新世中期に小笠原へ進入して種分化したものと推定された。このように本種の祖先における小笠原への進入について「いつ」と「どこから」は一応の解決を見た。しかし、「どのように」についてはまだはっきりと分からず、今後の課題となる。
オガサワラセセリの存続には、グリーンアノールやモクマオウ、リュウキュウマツ、ギンネムなどの外来生物が食草群落内で増加・拡大することを防ぐ手段を考えることが最重要課題として挙げられる。成虫の活動範囲は海岸部だけでなく、内陸部も広範に利用することから、これらの場所の保全も欠かすことができない。また、成虫は一年通して吸蜜植物が必要となることから、十分な量の吸蜜植物も確保しなければならない。母島では各地でグリーンアノールの防護柵を造設しつつあり(戸田ら, 2009)、柵内でグリーンアノール駆除を継続的に行っている場所では多くの幼虫等を観察することができた事例を経験している。さらに平島や向島などの母島属島では今のところグリーンアノールが見られないことから、父島の二の舞とならないように、何よりも現在の生息コアエリアである母島属島にこの外来トカゲを侵入させないことが必須であろう。耐震改修工事と今回のコロナ禍により本館展示場が開館できない状況が続いているが、常設展ではオガサワラセセリがオガサワラシジミとともに標本として展示(図7)されているので、展示場への入館が再開された際には、本稿で綴られた背景を思い出しながらご覧頂けると幸いである。
今回の戦前のオガサワラセセリ標本を筆頭とする多くの貴重な昆虫標本の移管に関して、(公財)山階鳥類研究所総裁の秋篠宮文仁皇嗣殿下、壬生基博理事長、奥野卓司所長、山崎剛史自然誌研究室長など、多くの方々にご支援を賜った。また、本稿作成にあたり王敏教授および范骁凌教授(華南農業大学)、千葉秀幸博士(ビショップ博物館)、Michael F. Braby准教授(オーストラリア国立大学)、中村康弘氏(日本チョウ類保全協会)、忠地良夫氏にもご協力頂いた。本研究はJSPS科研費(17K07528)の成果も一部含んでいる。関係者各位に心よりお礼を申し上げる。
引用文献
朝比奈英三, 1932. Zephyrus, 4 (2/3): 219.
Huang, Z., Chiba, H., Guo, D., Yago, M., Braby, M. F., Wang, M. and Fan, X., 2018. Mol. Phylogenet. Evol., 139: 106545.
石井良則, 2015. 小笠原研究年報 (38): 1―16.
加藤 真, 1991. 小笠原研究, (17/18): 32―59.
苅部治紀・高桑正敏・須田真一・松本浩一・岸本年郎・中原直子・長瀬博彦・鈴木瓦, 2004. 神奈川県立博物館調査研究報告(自然), (12): 65―86.
Matsumura, S., 1906. Annot. Zool. Japon., 6 (1): 7―15, pl. 1.
松村松年, 1906. 博物之友, (30): 13―16.
竹内誠一, 1936a. 昆蟲界, 4 (26): 264―272.
竹内誠一, 1936b. 昆蟲界, 4 (27): 351―353, pl. 145.
戸田光彦・中川直美・鋤柄直純, 2009. 地球環境, 14 (1): 36―46.
矢後勝也, 2014. 昆虫と自然, 49 (9): 4―7, 図巻頭1.
矢後勝也・須田真一・山崎剛史, 2018. 珠玉の昆虫標本-江戸から平成の昆虫研究を支えた東京大学秘蔵コレクション.東京大学出版会,東京.
矢後勝也, 中村康弘, 2007. Butterflies, (46): 35―45.
ウロボロスVolume25 Number1のトップページへ
海洋島で種分化したチョウの謎を解く
―小笠原固有のチョウ「オガサワラセセリ」―
矢後勝也(本館助教/昆虫自然史学・保全生物学)
本稿の主役・オガサワラセセリは年4~5回程度発生する多化性で、食草オガサワラススキ(イネ科)が繁茂する海岸線の草原帯で発生する(図2)。1905年に北海道帝国大学教授の松村松年(当時は札幌農学校教授)らにより父島で発見され、その翌年に松村自身の手により新種として発表されたのが最初である(Matsumura, 1906)。1935年には母島でも確認され(竹内, 1936b)、戦後になって母島属島の平島、向島、姉島からも記録された(加藤, 1991)。苅部ら(2004)により同じく母島属島の妹島、姪島からも報告がある。ただし父島からは戦前の記録しかない。また、戦後は1970年に母島で再発見されるまで長らく記録されなかった。そのためにオガサワラシジミが国の天然記念物に指定された1969年の時点で本種の生息が確認できず、文化庁による天然記念物選定の名簿から漏れた経緯があるが、生息範囲が狭いことからその希少性は高い。
このような本種であるが、実は最近になって北米原産の外来トカゲ・グリーンアノール(図3)の捕食圧により、これらのチョウ類をはじめとした多くの固有昆虫に絶滅の危機が迫っている(矢後・中村, 2007;矢後, 2014)。1960年年代に資材混入あるいはペットとして侵入したものがまず父島で増殖し、1984年に母島へ移入されて急増、さらに2013年には兄島でも多数繁殖していることが確認された。その生息数は現在500万頭とも600万頭とも推算されている。加えて海岸線ではモクマオウやリュウキュウマツ、ギンネム等の外来植物が、オガサワラセセリの食草オガサワラススキを被陰してその生長を阻害し、チョウ幼虫の成育に甚大な影響を及ぼしている(図4)。このため、環境省は2018年に種の保存法(絶滅のおそれのある野生動植物の種の保存に関する法律)の国内希少野生動植物種に指定し、捕獲・譲渡等を規制するとともに、母島やその属島での生息地の保全や保護増殖事業計画の策定等を行う準備を進めている。
一方、父島では戦後の記録がないことを述べたが、これはなぜだろうか?おそらく戦前の飛行場建設や築港の増加、宅地拡大、農地開発やそれに伴う農薬散布、軍の駐留による要塞化などで発生地のコアエリアが破壊されたことが主要因となり、残念ながら1940年代には絶滅したと考えられる。本種の存続には複数箇所の広大な海岸性草原が必要と思われ、台風や高波などによる塩害や季節的な変化、風向き、日照条件などから、時期ごとや年度ごとに幼生期の最適な生息場所や食草条件が少しずつ異なり、これに合わせて主要な発生場所を移動させているようである。要衝の中心であった父島やその属島では、そのような海岸性草原が真っ先に失われた可能性が高い。 父島からは1905年における本種最初の発見(Matsumura, 1906;松村, 1906)の他、戦前の朝比奈(1932)と竹内(1936b)による記録の計3例しか知られていない。しかも朝比奈の記録は伝聞、竹内の記録は目撃に留まり、これまで標本として存在するのは北海道大学に保管されている松村松年らのわずかな採集品のみであった。ところが、2017年冬に(公財)山階鳥類研究所から東大総合研究博物館に移管された昆虫標本の中から父島産オガサワラセセリの標本(図5)が見出されたのである(矢後ら, 2018)。ラベルには「Nov. 23, 1930. Chichijima, Ogasawara. K. Omichi leg.」と記載されている。標本の採集者K. Omichiは、小笠原島庁の勧業課長や植物検査官補としてミバエ類駆除の調査研究に従事していた大道金松と思われ(石井, 2015)、朝比奈(1932)の文中にも「ヲガサハラセセリ等に至つては島廳の大道氏の御話によると近年全く得られず、僅に同氏が本年の二月とかに二頭を得たに過ぎぬとの御話でした。」の記述が見られる。つまり大道がオガサワラセセリに精通し、実際に採集していたことは疑いない。故にこの標本データの信憑性は極めて高いことから、父島に間違いなく生息していたことを示す極めて重要な標本となるだろう。
ところで、小笠原のような海洋島でいつ、どこから、どのようにオガサワラセセリの祖先種が入り込み、固有種として分化したのか、という疑問が生じる。本種を含むセセリチョウ科のイチモンジセセリ属Parnaraは、アジアからオーストラリア、アフリカに分布し、世界からおよそ10種が知られ、日本にはオガサワラセセリP. ogasawarensisの他に、近縁種として本州以南に生息するイチモンジセセリP. guttataと南西諸島以南に分布するヒメイチモンジセセリP. badaの2種が生息する。特にオガサワラセセリはイチモンジセセリに最近縁とされていたものの、オーストラリアに分布するオセアニアイチモンジセセリP. amaliaに斑紋や♂交尾器がより類似することから、系統的位置の再検討も必要とされてきた。そこで筆者は中国の華南農業大学等との共同研究により、イチモンジセセリ属全種を用いて8つの遺伝子領域に基づいて分子系統解析を行い、分類学的再検討と系統地理学的考察を行った(Huang et al., 2019)。その結果が図6である。本属の共通祖先は漸新世中期に生じ、中新世後期から鮮新世後期にかけてアジア、オーストラリア、アフリカの各地域で分化を起こし、その後の分散と分断がアジアとアフリカでの多様化に大きな役割を果たしたと考えられた。ここで最も注目したいオガサワラセセリの系統的位置は、ヒマラヤ~中国、台湾、日本に広く生息するイチモンジセセリと中国南部~インドシナに分布するカクレイチモンジセセリP. battaで形成されるクレードの姉妹群として表されている。小笠原では稀にイチモンジセセリが記録されるが、オガサワラセセリはイチモンジセセリとは姉妹種にならないため、オガサワラセセリが独立種として維持されていることが理解できる。さらにオガサワラセセリの祖先種は北の日本本土方面や南のオセアニア方面の経由ではなく、むしろ琉球弧形成前の南西方面を経由して鮮新世中期に小笠原へ進入して種分化したものと推定された。このように本種の祖先における小笠原への進入について「いつ」と「どこから」は一応の解決を見た。しかし、「どのように」についてはまだはっきりと分からず、今後の課題となる。
オガサワラセセリの存続には、グリーンアノールやモクマオウ、リュウキュウマツ、ギンネムなどの外来生物が食草群落内で増加・拡大することを防ぐ手段を考えることが最重要課題として挙げられる。成虫の活動範囲は海岸部だけでなく、内陸部も広範に利用することから、これらの場所の保全も欠かすことができない。また、成虫は一年通して吸蜜植物が必要となることから、十分な量の吸蜜植物も確保しなければならない。母島では各地でグリーンアノールの防護柵を造設しつつあり(戸田ら, 2009)、柵内でグリーンアノール駆除を継続的に行っている場所では多くの幼虫等を観察することができた事例を経験している。さらに平島や向島などの母島属島では今のところグリーンアノールが見られないことから、父島の二の舞とならないように、何よりも現在の生息コアエリアである母島属島にこの外来トカゲを侵入させないことが必須であろう。耐震改修工事と今回のコロナ禍により本館展示場が開館できない状況が続いているが、常設展ではオガサワラセセリがオガサワラシジミとともに標本として展示(図7)されているので、展示場への入館が再開された際には、本稿で綴られた背景を思い出しながらご覧頂けると幸いである。
今回の戦前のオガサワラセセリ標本を筆頭とする多くの貴重な昆虫標本の移管に関して、(公財)山階鳥類研究所総裁の秋篠宮文仁皇嗣殿下、壬生基博理事長、奥野卓司所長、山崎剛史自然誌研究室長など、多くの方々にご支援を賜った。また、本稿作成にあたり王敏教授および范骁凌教授(華南農業大学)、千葉秀幸博士(ビショップ博物館)、Michael F. Braby准教授(オーストラリア国立大学)、中村康弘氏(日本チョウ類保全協会)、忠地良夫氏にもご協力頂いた。本研究はJSPS科研費(17K07528)の成果も一部含んでいる。関係者各位に心よりお礼を申し上げる。
引用文献
朝比奈英三, 1932. Zephyrus, 4 (2/3): 219.
Huang, Z., Chiba, H., Guo, D., Yago, M., Braby, M. F., Wang, M. and Fan, X., 2018. Mol. Phylogenet. Evol., 139: 106545.
石井良則, 2015. 小笠原研究年報 (38): 1―16.
加藤 真, 1991. 小笠原研究, (17/18): 32―59.
苅部治紀・高桑正敏・須田真一・松本浩一・岸本年郎・中原直子・長瀬博彦・鈴木瓦, 2004. 神奈川県立博物館調査研究報告(自然), (12): 65―86.
Matsumura, S., 1906. Annot. Zool. Japon., 6 (1): 7―15, pl. 1.
松村松年, 1906. 博物之友, (30): 13―16.
竹内誠一, 1936a. 昆蟲界, 4 (26): 264―272.
竹内誠一, 1936b. 昆蟲界, 4 (27): 351―353, pl. 145.
戸田光彦・中川直美・鋤柄直純, 2009. 地球環境, 14 (1): 36―46.
矢後勝也, 2014. 昆虫と自然, 49 (9): 4―7, 図巻頭1.
矢後勝也・須田真一・山崎剛史, 2018. 珠玉の昆虫標本-江戸から平成の昆虫研究を支えた東京大学秘蔵コレクション.東京大学出版会,東京.
矢後勝也, 中村康弘, 2007. Butterflies, (46): 35―45.
ウロボロスVolume25 Number1のトップページへ

図1 オガサワラススキに止まるオガサワラセセリ♀.前翅長は20 mm前後.

図2 オガサワラセセリの生息環境(母島南崎).

図3 北米原産の外来トカゲ・グリーンアノール.

図4 海岸線における草原面積の変化(母島南崎).a: 1956年6月撮影の空中写真.b: 2013年12月撮影の空中写真.海岸線の白く見える部分がオガサワラススキなどで構成された海岸性草原.aと比較してbの海岸性草原はかなり減退していることが分かる.国土地理院ウェブサイト(https://mapps.gsi.go.jp/maplibSearch.do#1)より.

図5 (公財)山階鳥類研究所旧蔵の父島産オガサワラセセリ♂標本.
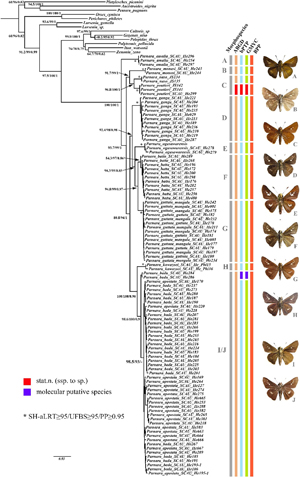
図6 8遺伝子領域を用いて最尤法により得られたイチモンジセセリ属の分子系統樹(Mol. Phylogenet. Evol., 139: 106545より).Eがオガサワラセセリを示す.

図7 本館常設展で展示されているオガサワラセセリとオガサワラシジミの標本.