
小石川分館特別展示
貝殻の螺旋と数理モデル
化石が主な研究対象である古生物学という学問では、現代に生きている生物を扱う他の学問と比べて、研究対象から得られる情報が極めて限られる。化石からは行動や生体分子などの情報が大きく損なわれるため、形態から得られる情報が研究において主要な役割を果たす。そのような理由もあり、生物の形態解析手法の今日までの発展は古生物学によるところが大きい。その中で、理論形態学という分野において、貝殻の形を数理モデルで再現する試みがなされてきた。ひとつ、留意点として、理論形態学は、形を数理モデルで表すことが目的なのではなく、生物の生態や多様性、進化を明らかにすることが目的である。形を数理モデルで近似することができると、モデルのパラメーターを調整することで、様々な種類の貝殻の形はもちろん、現実には存在しない形をつくることができる。このような数理モデルから理論的に想定される様々な形(理論形態)の中から、現在生きているような貝殻の形がなぜ選ばれたかを探る学問が理論形態学である。ここからは、理論形態学の発展とともにつくられてきた、貝殻の形を表現する数理モデルを紹介していく。理論形態学の手法として、生物の生態や多様性、進化を明らかにするためのいくつかの理論的枠組みがあり、数理モデルと密接な関係があるのだが、ここでは深くは立ち入らず、あえて「貝殻の形を数理モデルで表現すること」そのものに焦点を当てていく。貝殻の形の数理的な美しさを感じていただければ幸いである。
等角螺旋
巻貝やオウムガイ、アンモナイトの貝殻を見ると、規則的な美しい螺旋でできていることがわかるだろう。この螺旋は「等角螺旋」あるいは「対数螺旋」と呼ばれる(図1, 2)。巻き軸に垂直な直線を引いたときに、直線と螺旋が一定の角度で交わることから「等角螺旋」、巻きの軸からの距離と巻きの数の関係を数式に表すと対数関数が現れることから「対数螺旋」である。貝殻の螺旋が等角螺旋で近似できることは古くから言われていたが、D'Arcy W. Thompsonによる1917年の著作、“On Growth and Form”(邦題「生物のかたち」)で詳しくまとめられた。等角螺旋の特徴は、螺旋の形がスケールに依存しないことである。螺旋を縮小あるいは拡大しても螺旋の見た目はまったく変化しない。
さて、貝殻の形を測って等角螺旋を表す数式で書き表すことを考えよう。直感的には、まずは巻き軸を特定し、軸からの距離と巻きの数をいくつかの点で測り、それを適切に再現できるようなパラメーターを選べばよい。しかし実は、対角螺旋の式のパラメーターを推定するためには、巻きの軸の位置は必ずしも必要ではない。先ほど述べたように対角螺旋の形はスケールに依存しないので、オウムガイのような平面巻きでは本質的なパラメーターは1つしかなく、その1つのパラメーターを推定するためには、ある部位の大きさが1巻の間に何倍になるかがわかればよい。この考え方は後述するRaupのモデルにつながる。
次に、等角螺旋を描いて貝殻の形を表現することを考えよう。等角螺旋の式で表されるのは一本の線に過ぎないので、貝殻の形をつくるには何かしらの工夫が必要である。例えば、螺旋を中心としたチューブを考える、あるいは、貝殻表面を螺旋の線の集合として表すような方法がある。対角螺旋は、それ単体では貝殻の形を近似するモデルというより、貝殻の形の特徴の本質を記述したものといったほうがよいのかもしれない。
Raupモデル
アメリカの古生物学者David M. Raupは、コンピューターグラフィックスを用いることで、数理モデルが表現する形とその多様性を効果的に視覚化することに成功した。母曲線と呼ばれる閉曲線、つまり円や楕円などの閉じた曲線が、等角螺旋に沿って拡大しながら動くとき、その軌跡は貝殻の形をよく近似する。Raupは、この数理モデルを用いて形の多様性に関する解析を行った (Raup, 1966)。Raupモデルは、パラメーター3つで貝殻の形を表現する(図3, 4)。3つのパラメーターは具体的には以下の通りである。(1) 拡張率W:巻きが1周したときに大きさが何倍になるか。(2) 母曲線の位置D:母曲線が巻き軸からどれだけ離れているか。(3) 転移率T:巻き軸からの距離に対してどのくらい巻き軸方向に成長するか。直感的には3つのパラメーターはそれぞれ、巻きの緩さ、臍の大きさ、殻頂の尖り具合を表す。これらのパラメーターを変化させることで、アンモナイトや一般的な巻貝、笠形の貝、二枚貝など多くの貝殻形を表現することができる。
Raupモデルのパラメーターは3つとも、2つの部位の長さの比という形で表される。長ささえ測定できれば、実際の貝殻をRaupモデルのパラメーターで容易に表すことができる。Tに関わる測定のためには母曲線の中心を推定しなければならないので、多少の工夫は必要であるが、一般的な形の巻貝であればそれほど問題なくパラメーターの推定ができるだろう。ただし、笠形の貝や二枚貝は巻きが1周しない場合がほとんどなのでパラメーターの推定は非常に困難である。現在はX線CT装置を使用することで、貝殻の透過断面や3Dデータをつくることができるので、パラメーター推定をより容易に正確に行うことができる。また、実際に貝殻を切断することでも必要なデータを得ることが出来る。 Raupモデルから貝殻の形を再現するには、まず母曲線、すなわち殻口の形を決める必要がある。円や楕円で近似できるものは簡単だが、複雑な形のものは座標上にその形をうまくプロットしなくてはならないので少し面倒である。最初の母曲線をDを満たすようにプロットし、WとTをもとに等角螺旋に沿って描写することでモデル通りの形をつくることができる。さらに、RaupはTを巻きの途中で連続的に変化させることで、巻貝によく見られる比較的変則的な巻きも再現した。
当時、Raupモデルは貝殻の形を視覚的に再現する画期的な数理モデルであったが、近年は表現力の高い他のモデルが存在することもあり、形を再現するときには他のモデルが使われることも多い。しかし、パラメーターの計測の容易さはRaupモデルの強みであり、現在でも実際の貝殻の形を計測値から数理モデルに書き表すときには、まず候補に挙がるモデルである。
成長管モデル
成長管モデル(Okamoto, 1988)は、異常巻アンモナイトと呼ばれる通常の螺旋とはかけ離れた巻き方のアンモナイトを説明するためにつくられたモデルである(図6)。再現力が高いことがこのモデルの特徴で、なめらかな管状のものであればどんな形をしていても再現が可能である。貝殻の各成長段階で、母曲線の「拡大」、成長の「曲がり」「捩れ」を考えることで形を表現する(図5, 7)。Raupモデルでつくられるような対数螺旋は成長管モデルでつくられる最も単純な形であり、成長管モデルを構成する3つのパラメーター、螺管拡大率E、規格化曲率C、規格化捩率Tを一定に保ったときにつくられる。成長管モデルの巧みな点のひとつは、3つのパラメーターを母曲線の大きさで規格化しているところである。これによりスケールへの依存性がなくなり、単純なパラメーターで等角螺旋をつくることができる。また生物の発生として考えると、母曲線と同じスケールで成長する軟体部の視点で見たときに自然なパラメーターとなることも規格化による恩恵である。もうひとつ、Raupモデルとは異なり、あらかじめ巻き軸を設定しないことも特徴であり、生物の発生で自然に螺旋が形成されることが示唆される。
実際の貝殻から成長管モデルのパラメーターを直接測定するのは非常に困難である。現実的には、ほとんど同じ形をつくることのできるRaupモデルで表現した後、パラメーターを変換する計算により成長管モデルのパラメーターで表現し直すことが無難な方法だろう。対数螺旋から外れた形については、さらに込み入った方法でパラメーター推定を行わなければならない。
既に決定したパラメーターから貝殻をコンピューターグラフィックスで再現するとき、成長管モデルの真価が発揮される。母曲線を拡大させながら移動させていくことで貝殻を再現することはRaupモデルと同じであるが、Raupモデルはあらかじめ定めた巻き軸を基準にD、W、Tから毎度位置を計算する必要があるのに対し、成長管モデルは最初に母曲線の位置と成長方向だけ定めておけば母曲線自身を基準として次々と計算していくことができる。さらに、3つのパラメーターE、C、Tは各成長段階で定義される成長に関わる量であり、母曲線の移動とパラメーターの関係性が直感的にわかりやすい。また、成長とともにパラメーターが変化する場合もRaupモデルに比べて自然に表現することができる。
成長管モデルは、その高い表現力から、複雑な形を再現するための発生的、生態的な条件を調べる目的に適している。Raupモデルとは異なり、成長管モデルは実際の貝殻からパラメーターを測定することは困難であるが、そのパラメーターには発生学的な意味を見出すことが可能であり、形の形成を説明するモデルとして非常に優れている。
今回紹介した以外にも様々な数理モデルが存在するが、貝殻の基本的な形を表現するモデルとしては上述のモデルでほとんど完成されていると言ってよい。これからの時代に求められるモデルは、形を表現するものではなく、発生における化学的、力学的なプロセスから形の形成を説明するようなモデルであろう。
ウロボロスVolume24 Number2のトップページへ
貝殻の螺旋と数理モデル
佐藤英明(理学系研究科修士課程/古生物学)
化石が主な研究対象である古生物学という学問では、現代に生きている生物を扱う他の学問と比べて、研究対象から得られる情報が極めて限られる。化石からは行動や生体分子などの情報が大きく損なわれるため、形態から得られる情報が研究において主要な役割を果たす。そのような理由もあり、生物の形態解析手法の今日までの発展は古生物学によるところが大きい。その中で、理論形態学という分野において、貝殻の形を数理モデルで再現する試みがなされてきた。ひとつ、留意点として、理論形態学は、形を数理モデルで表すことが目的なのではなく、生物の生態や多様性、進化を明らかにすることが目的である。形を数理モデルで近似することができると、モデルのパラメーターを調整することで、様々な種類の貝殻の形はもちろん、現実には存在しない形をつくることができる。このような数理モデルから理論的に想定される様々な形(理論形態)の中から、現在生きているような貝殻の形がなぜ選ばれたかを探る学問が理論形態学である。ここからは、理論形態学の発展とともにつくられてきた、貝殻の形を表現する数理モデルを紹介していく。理論形態学の手法として、生物の生態や多様性、進化を明らかにするためのいくつかの理論的枠組みがあり、数理モデルと密接な関係があるのだが、ここでは深くは立ち入らず、あえて「貝殻の形を数理モデルで表現すること」そのものに焦点を当てていく。貝殻の形の数理的な美しさを感じていただければ幸いである。
等角螺旋
巻貝やオウムガイ、アンモナイトの貝殻を見ると、規則的な美しい螺旋でできていることがわかるだろう。この螺旋は「等角螺旋」あるいは「対数螺旋」と呼ばれる(図1, 2)。巻き軸に垂直な直線を引いたときに、直線と螺旋が一定の角度で交わることから「等角螺旋」、巻きの軸からの距離と巻きの数の関係を数式に表すと対数関数が現れることから「対数螺旋」である。貝殻の螺旋が等角螺旋で近似できることは古くから言われていたが、D'Arcy W. Thompsonによる1917年の著作、“On Growth and Form”(邦題「生物のかたち」)で詳しくまとめられた。等角螺旋の特徴は、螺旋の形がスケールに依存しないことである。螺旋を縮小あるいは拡大しても螺旋の見た目はまったく変化しない。
さて、貝殻の形を測って等角螺旋を表す数式で書き表すことを考えよう。直感的には、まずは巻き軸を特定し、軸からの距離と巻きの数をいくつかの点で測り、それを適切に再現できるようなパラメーターを選べばよい。しかし実は、対角螺旋の式のパラメーターを推定するためには、巻きの軸の位置は必ずしも必要ではない。先ほど述べたように対角螺旋の形はスケールに依存しないので、オウムガイのような平面巻きでは本質的なパラメーターは1つしかなく、その1つのパラメーターを推定するためには、ある部位の大きさが1巻の間に何倍になるかがわかればよい。この考え方は後述するRaupのモデルにつながる。
次に、等角螺旋を描いて貝殻の形を表現することを考えよう。等角螺旋の式で表されるのは一本の線に過ぎないので、貝殻の形をつくるには何かしらの工夫が必要である。例えば、螺旋を中心としたチューブを考える、あるいは、貝殻表面を螺旋の線の集合として表すような方法がある。対角螺旋は、それ単体では貝殻の形を近似するモデルというより、貝殻の形の特徴の本質を記述したものといったほうがよいのかもしれない。
Raupモデル
アメリカの古生物学者David M. Raupは、コンピューターグラフィックスを用いることで、数理モデルが表現する形とその多様性を効果的に視覚化することに成功した。母曲線と呼ばれる閉曲線、つまり円や楕円などの閉じた曲線が、等角螺旋に沿って拡大しながら動くとき、その軌跡は貝殻の形をよく近似する。Raupは、この数理モデルを用いて形の多様性に関する解析を行った (Raup, 1966)。Raupモデルは、パラメーター3つで貝殻の形を表現する(図3, 4)。3つのパラメーターは具体的には以下の通りである。(1) 拡張率W:巻きが1周したときに大きさが何倍になるか。(2) 母曲線の位置D:母曲線が巻き軸からどれだけ離れているか。(3) 転移率T:巻き軸からの距離に対してどのくらい巻き軸方向に成長するか。直感的には3つのパラメーターはそれぞれ、巻きの緩さ、臍の大きさ、殻頂の尖り具合を表す。これらのパラメーターを変化させることで、アンモナイトや一般的な巻貝、笠形の貝、二枚貝など多くの貝殻形を表現することができる。
Raupモデルのパラメーターは3つとも、2つの部位の長さの比という形で表される。長ささえ測定できれば、実際の貝殻をRaupモデルのパラメーターで容易に表すことができる。Tに関わる測定のためには母曲線の中心を推定しなければならないので、多少の工夫は必要であるが、一般的な形の巻貝であればそれほど問題なくパラメーターの推定ができるだろう。ただし、笠形の貝や二枚貝は巻きが1周しない場合がほとんどなのでパラメーターの推定は非常に困難である。現在はX線CT装置を使用することで、貝殻の透過断面や3Dデータをつくることができるので、パラメーター推定をより容易に正確に行うことができる。また、実際に貝殻を切断することでも必要なデータを得ることが出来る。 Raupモデルから貝殻の形を再現するには、まず母曲線、すなわち殻口の形を決める必要がある。円や楕円で近似できるものは簡単だが、複雑な形のものは座標上にその形をうまくプロットしなくてはならないので少し面倒である。最初の母曲線をDを満たすようにプロットし、WとTをもとに等角螺旋に沿って描写することでモデル通りの形をつくることができる。さらに、RaupはTを巻きの途中で連続的に変化させることで、巻貝によく見られる比較的変則的な巻きも再現した。
当時、Raupモデルは貝殻の形を視覚的に再現する画期的な数理モデルであったが、近年は表現力の高い他のモデルが存在することもあり、形を再現するときには他のモデルが使われることも多い。しかし、パラメーターの計測の容易さはRaupモデルの強みであり、現在でも実際の貝殻の形を計測値から数理モデルに書き表すときには、まず候補に挙がるモデルである。
成長管モデル
成長管モデル(Okamoto, 1988)は、異常巻アンモナイトと呼ばれる通常の螺旋とはかけ離れた巻き方のアンモナイトを説明するためにつくられたモデルである(図6)。再現力が高いことがこのモデルの特徴で、なめらかな管状のものであればどんな形をしていても再現が可能である。貝殻の各成長段階で、母曲線の「拡大」、成長の「曲がり」「捩れ」を考えることで形を表現する(図5, 7)。Raupモデルでつくられるような対数螺旋は成長管モデルでつくられる最も単純な形であり、成長管モデルを構成する3つのパラメーター、螺管拡大率E、規格化曲率C、規格化捩率Tを一定に保ったときにつくられる。成長管モデルの巧みな点のひとつは、3つのパラメーターを母曲線の大きさで規格化しているところである。これによりスケールへの依存性がなくなり、単純なパラメーターで等角螺旋をつくることができる。また生物の発生として考えると、母曲線と同じスケールで成長する軟体部の視点で見たときに自然なパラメーターとなることも規格化による恩恵である。もうひとつ、Raupモデルとは異なり、あらかじめ巻き軸を設定しないことも特徴であり、生物の発生で自然に螺旋が形成されることが示唆される。
実際の貝殻から成長管モデルのパラメーターを直接測定するのは非常に困難である。現実的には、ほとんど同じ形をつくることのできるRaupモデルで表現した後、パラメーターを変換する計算により成長管モデルのパラメーターで表現し直すことが無難な方法だろう。対数螺旋から外れた形については、さらに込み入った方法でパラメーター推定を行わなければならない。
既に決定したパラメーターから貝殻をコンピューターグラフィックスで再現するとき、成長管モデルの真価が発揮される。母曲線を拡大させながら移動させていくことで貝殻を再現することはRaupモデルと同じであるが、Raupモデルはあらかじめ定めた巻き軸を基準にD、W、Tから毎度位置を計算する必要があるのに対し、成長管モデルは最初に母曲線の位置と成長方向だけ定めておけば母曲線自身を基準として次々と計算していくことができる。さらに、3つのパラメーターE、C、Tは各成長段階で定義される成長に関わる量であり、母曲線の移動とパラメーターの関係性が直感的にわかりやすい。また、成長とともにパラメーターが変化する場合もRaupモデルに比べて自然に表現することができる。
成長管モデルは、その高い表現力から、複雑な形を再現するための発生的、生態的な条件を調べる目的に適している。Raupモデルとは異なり、成長管モデルは実際の貝殻からパラメーターを測定することは困難であるが、そのパラメーターには発生学的な意味を見出すことが可能であり、形の形成を説明するモデルとして非常に優れている。
今回紹介した以外にも様々な数理モデルが存在するが、貝殻の基本的な形を表現するモデルとしては上述のモデルでほとんど完成されていると言ってよい。これからの時代に求められるモデルは、形を表現するものではなく、発生における化学的、力学的なプロセスから形の形成を説明するようなモデルであろう。
ウロボロスVolume24 Number2のトップページへ
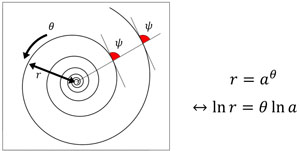
図1 等角螺旋.中心から引いた直線と螺旋のなす角が常に一定となる.オウムガイやアンモナイトの形をおおよそ近似する.
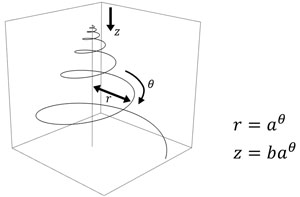
図2 立体的な等角螺旋.真横から見ると円錐形をしており,巻貝の形に似ている.
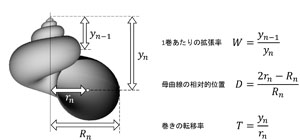
図3 Raupモデルを構成するパラメーター.実際の貝殻から直接計測が可能である.Tは研究によって複数の定義があるが,ここではRaup (1966)の定義によった.
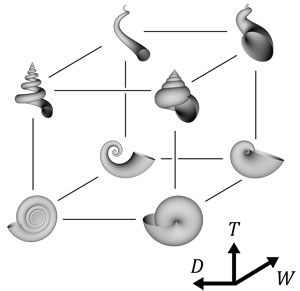
図4 Raupモデルの3つのパラメーターを変化させて描くことのできる様々な形.Wは巻きの緩さ,Dは臍の広さ,Tは殻頂の尖り具合である.このように,各パラメーターを軸とした空間を形態空間といい,空間中のそれぞれの点に対応する形が存在する.Raup (1966)は現実の貝殻に見られる形が形態空間内で分布するパターンを見出した.
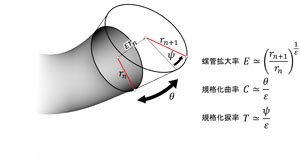
図5 成長環モデルを構成するパラメーター.厳密には各パラメーターは微分によって得られるが,微小な成長を考えることで近似的に考えることができる.

図6 成長管モデルで表現することのできる異常巻アンモナイトNipponites mirabilis.Okamoto (1988) はアンモナイトの生息姿勢にある規則を課すことで,自然にこの複雑な形が再現されることを発見した.
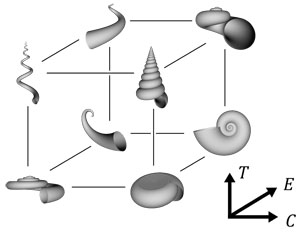
図7 成長管モデルの3つのパラメーターを変化させて描くことのできる様々な形.Eは拡大率,Cは巻きの強さ,Tは捩れの強さである.